The short- and long-term effects of chronic ketamine during adolescence on object recognition memory in rats
For my master’s thesis, I was interested in how chronic drug abuse during late adolescence could impair cognition in not only the short-term, but also if impairments could persist into adulthood. At the most basic level, I was interested in seeing if ketamine, an anesthetic commonly abused for its hallucinogenic properties, abuse during this critical period of development would impair spatial and non-spatial object recognition memory.
Two main questions I sought to answer:
- Does chronic ketamine interact with typical neurodevelopmental process to induce cognitive/behavioral alterations that show not only immediate effects but also effects that persist into adulthood? Are the effects enhanced in adulthood.
- Does chronic subanesthetic ketamine administration during adolescence differentially affect spatial and non-spatial recognition memory?
To achieve this, I chronically administered ketamine (a subanesthetic dosage once daily for seven consecutive days) in rats during adolescence and tested them on two behavioral tasks, the object recognition (ORT) and object location (OLT) tasks, which test non-spatial and spatial recognition memory, respectively.
Behavioral tasks:
Object Recognition Task (ORT)
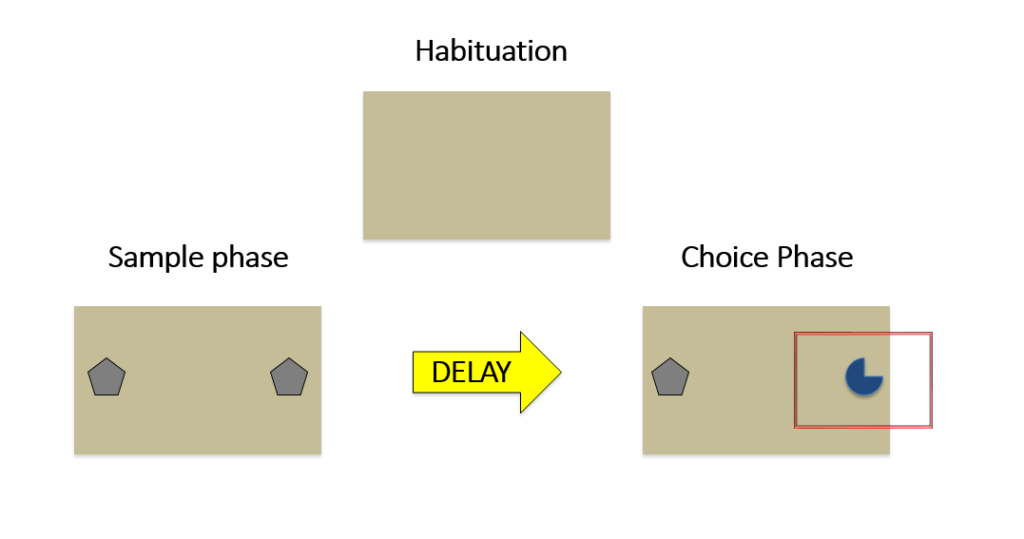
The typical design for the object recognition task (ORT) takes place in an open box and begins with an habituation phase. Following the drug administration schedule, rats underwent habituation, where they were placed in the open box, without objects, and allowed to freely explore for 10 minutes. Of course, this time can vary based on experimental questions; however, prior research had used a similar paradigm when investigating the effects of ketamine on the ORT. Following three consecutive days of habituation, the rats were subjected to the sample (study) phase, where two identical objects were placed in the box and they were allowed to freely explore for 3 minutes. Then, after 15 minutes and 24 hours (to test short- and long-term recognition memory, respectively), the rats underwent choice phases. In these phases, one of the familiar objects from the sample phase is presented, along with a novel object in which the rat had never experienced. The objects used as the novel objects during the 15-minute and 24-hour choice phases were different, ensuring that the objects were truly novel upon presentation to rats.
Object Location Task (OLT)
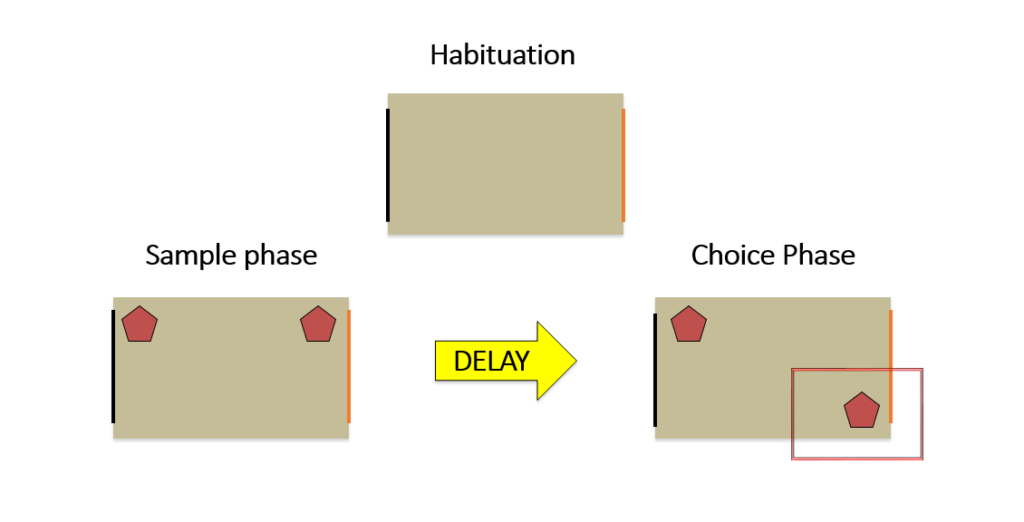
Similar in design to the ORT, the object location task (OLT) takes place in an open box and begins with an habituation phase, followed by sample and choice phases. However, while the ORT tests non-spatial recognition memory, the OLT is more a test of spatial recognition memory and has been shown to induce activity in different brain regions than the ORT. Rather than presenting novel objects, two identical objects are used throughout all of the phases but moved to novel locations within the box. Again, the rats underwent three days of 10-minute habituation phases, followed by a sample phase on the fourth day and a choice phase 15 minutes and 24 hours following the sample phase. The locations used as the novel locations during the 15-minute and 24-hour choice phases were different, ensuring that the locations were truly novel upon presentation to the rats.
In both the ORT and OLT, the main measurements were total exploration time of the objects/locations in seconds as well as a discrimination ratio, which is simply a ratio of the difference in time spent exploring the novel and familiar objects/locations divided by the total exploration time.
Main findings:
Ketamine decreased total exploration and time exploring novel object during ORT
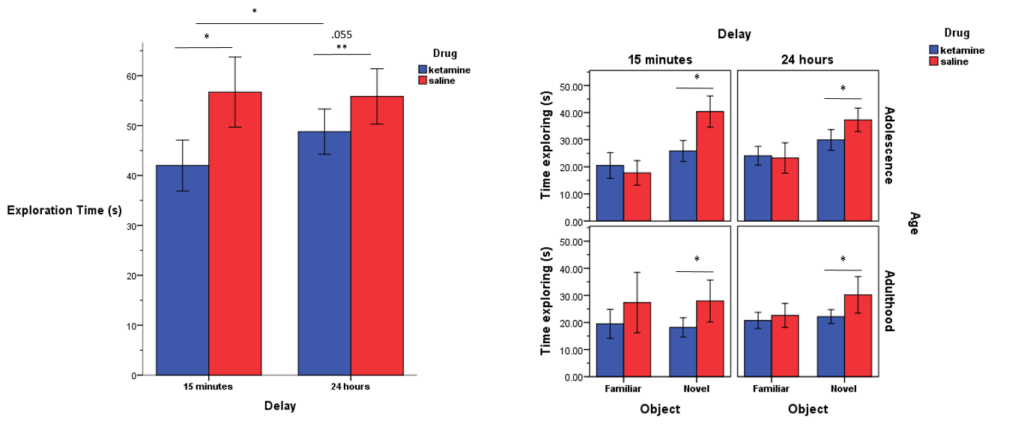
The figure on the left represents the total exploration time of the drug groups (i.e., ketamine and saline) during both choice-phase delays (i.e., 15 minutes and 24 hours). Not only did ketamine-treated rats explore significantly less overall, on average, compared to the saline-treated rats during both 15-minute and 24-hour choice phases, but there were differences within the ketamine-treated group too, as they explored the objects less during the 15-minute choice phase compared to the 24-hour choice phase, potentially indicating that ketamine might have caused the rats to take longer to properly habituate to the task.
The figure on the right demonstrates the differences in exploration time of the familiar vs novel objects during both 15-minute and 24-hour choice phases during both adolescence and adulthood. Ketamine-treated rats spent less time exploring the novel objects during both choice phases (i.e., 15 minutes and 24 hours) at both age points (i.e., adolescence and adulthood)., suggesting both immediate and long-lasting deficits in object recognition memory induced by chronic ketamine administration during adolescence.
Ketamine impaired memory for the familiar object
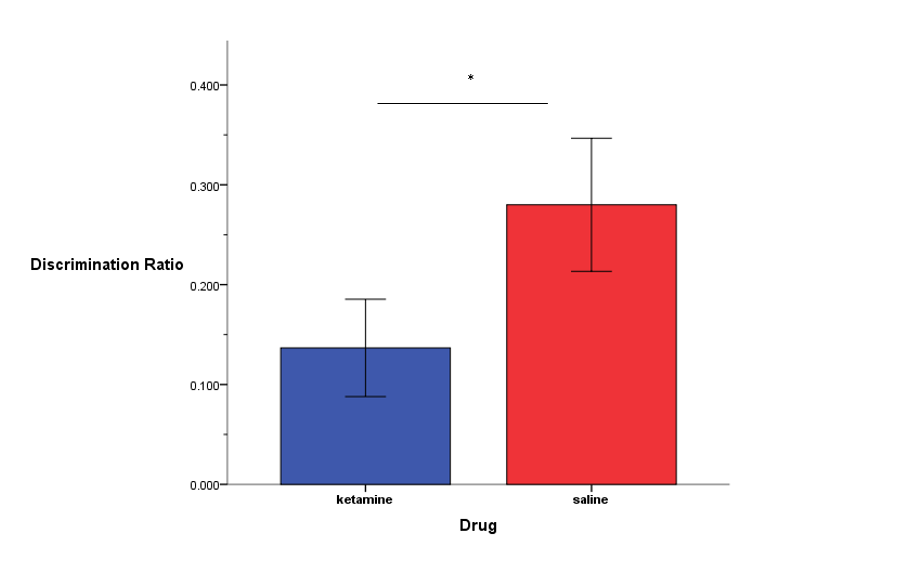
The discrimination ratio, which, again, is the ratio of the difference in time spent exploring the novel and familiar objects divided by the total exploration time, was significantly lower in ketamine-treated rats, suggesting impaired memory for the familiar object during the ORT.
Ketamine did not decrease total exploration or time exploring novel location during OLT
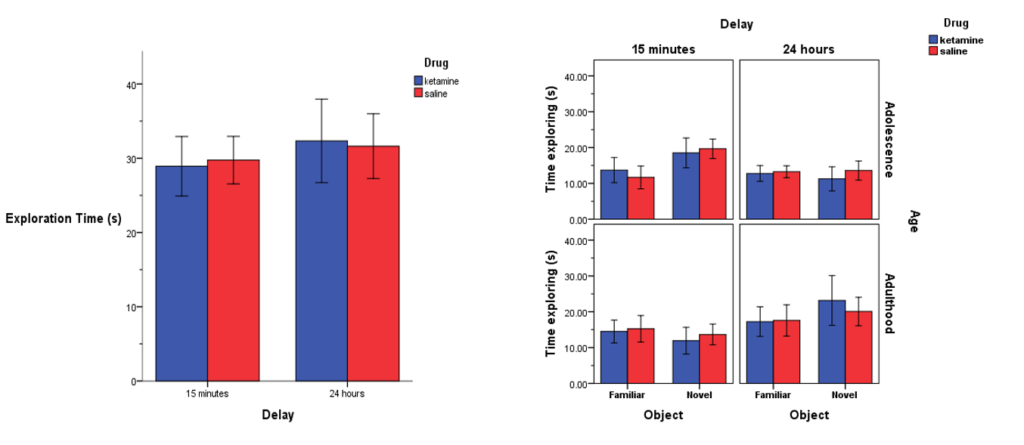
Unlike the ORT, there were no differences in total exploration time of the locations during the OLT, which can be seen in the figure on the left.
Additionally, as seen in the figure on the right, there were also no differences in time spent exploring the familiar vs novel locations during either choice phase (i.e., 15 minutes and 24 hours) or age point (i.e., adolescence and adulthood).
Ketamine impaired memory for the familiar location
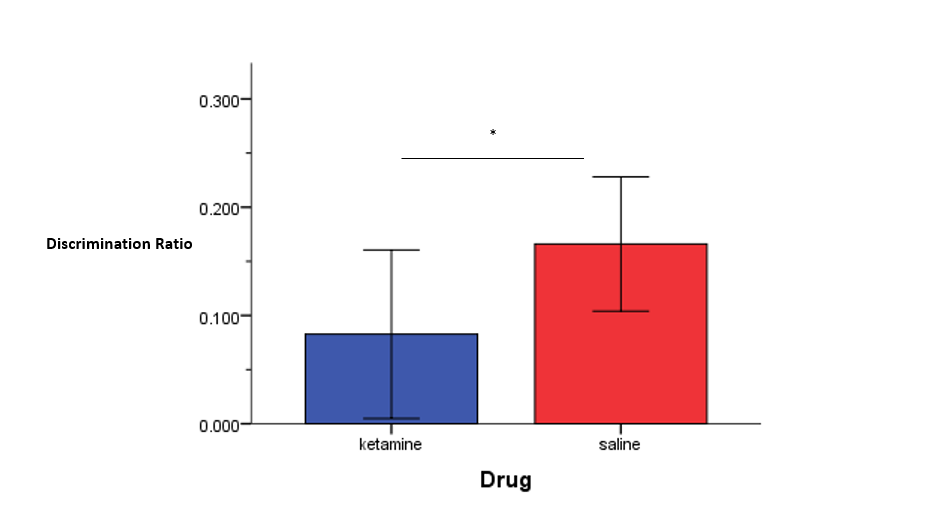
However, although there were no differences in total exploration time between the ketamine- and saline-treated rats during the OLT, the ketamine-treated rats, on average, had significantly lower discrimination ratios compared to the saline-treated rats, indicating that the ketamine-treated did indeed have impaired memory for the familiar location during the OLT. This effect is similar to the effect found in the ORT, although not as robust.
Summary
- Chronic ketamine during adolescence impairs short- and long-term object recognition memory during both adolescence and adulthood
- Effects of ketamine on ORT more robust, indicating a stronger effect on non-spatial object recognition memory.
- Persisting effects of ketamine are seen in adulthood
What does this mean for humans?
Prior research has shown that ketamine can induce symptoms similar to the positive, negative, and cognitive symptoms of schizophrenia. The ORT and OLT are considered to contain an episodic memory-like component, which is impaired in individuals with schizophrenia. Additionally, there is a lot of emerging evidence pointing toward schizophrenia as a neurodevelopmental illness and NMDA hypofunction is considered as a convergence point for the progression and symptoms of it. Thus, being that chronic administration of NMDA antagonists, like ketamine, mimics the symptoms seen in this devastating illness, this line of research may provide novel ways to investigate potential causes, mechanisms, and interventions through multidisciplinary approaches.
For more on this research, as well as statistics and a full discussion of the findings, check the original paper, which can be found in its entirety here.
Following graduate school, I continued to work in a behavioral neuroscience lab studying basic mechanisms underlying mental illnesses; specifically, the lab investigated electrophysiological mechanisms underlying genetic models of anxiety and schizophrenia. I was involved in many different projects while there and some of that work can be seen below.
Presentations, Proceedings, and Publications
Hardin WD, Hunter AS. (2015, March). Chronic ketamine during adolescence impairs object recognition memory in rats. Poster session presented at the annual meeting of the Eastern Psychological Association, New York, NY.
, , , , , , , , , , , ,
Padilla-Coreano N, Bolkan SS, Pierce GM, Blackman DR, Hardin WD, Garcia-Garcia AL, Spellman TJ, Gordon JA. Direct ventral hippocampal-prefrontal input is required for anxiety-related neural activity and behavior. Neuron 89(4): 857-66. 2016.